
S otázkou dynamiky pôvodných pralesov súvisí otázka vývojových štádií. Teoreticky existujú dva typy dynamiky pralesa – malý a veľký vývojový cyklus. V malom cykle, považovanom za typický pre lesy mierneho pásma, sa v priebehu storočí striedajú tri vývojové štádiá: štádium dorastania, štádium optima a štádium rozpadu. Tieto tri štádiá sa striedajú na malých plochách, takže pralesovité porasty v celkovom pohľade zostávajú takmer nemenné. Odumieranie stromov v štádiu rozpadu býva spôsobené ich vekom a nikdy nebýva masové. Niektoré zo starých stromov vždy prežívajú do nového štádia dorastania, prípadne aj dlhšie. Klimaxové porasty sú spravidla tvorené tiennymi drevinami, napr. smrekom, bukom a jedľou, len na extrémnych stanovištiach sa v klimaxe udržiavajú slnné dreviny ako smrekovec, borovica či pionierske dreviny.
U severských (boreálnych) lesov býva vývoj odlišný. Vďaka chladnej klíme a zamokreným pôdam tu dochádza k hromadeniu humusu, ktorý postupne znemožní prirodzenú obnovu drevín. Les sa preto nakoniec rozpadá (starnutím a rednutím porastov) a umožní rozklad humusu. Tento proces je spravidla urýchlený vetrovými kalamitami a požiarmi. Na takto uvoľnených plochách vznikajú porasty pionierskych drevín (breza, osika, borovica), ktorý je postupne podrastený smrekom, mení sa na smrečiny s výrazným hromadením humusu a veľký vývojový cyklus sa opakuje.
Vzťah medzi malým a veľkým vývojovým cyklom sa dá vyjadriť nasledovne:
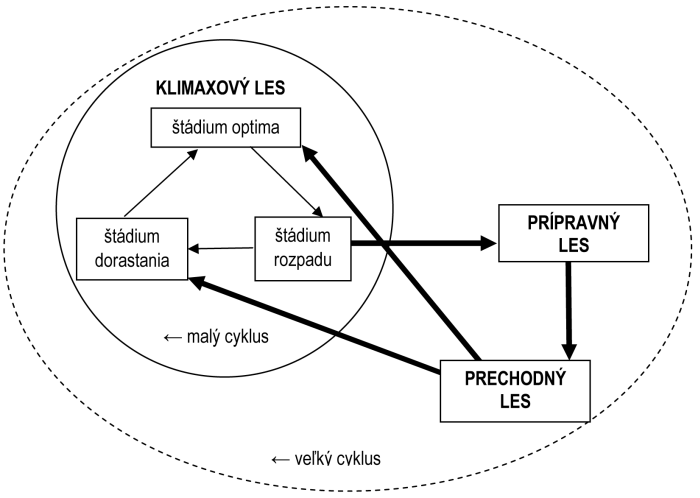
Veľký a malý vývojový cyklus lesa
Podľa (MÍCHAL a kol., 1992)
Odvolávanie sa na veľký vývojový cyklus sa v poslednej dobe stalo medzi ochranármi módou. Najmä v prípade odumierania smrečín sa často stretávame s názormi, že ich rozpad vplyvom vetra a podkôrneho hmyzu, rovnako ako ich veľkoplošná premena na porasty jarabiny, či aspoň premena starých smrečín na mladiny, sú len prirodzenou súčasťou ich vývoja a netreba sa nimi znepokojovať. Tieto názory majú za cieľ obhájiť, aby sa do chránených území nezasahovalo ani v prípade masového odumierania smreka. Ide však o názory, ktoré sú do značnej miery účelové. V súčasnosti sa všeobecne sa predpokladá, že „odbočky“ z malého do veľkého vývojového cyklu sú v miernom pásme zriedkavé [5] a že smrečiny mierneho pásma sa udržiavajú bez dramatických zmien drevinového zloženia a štruktúry.
Dá sa, samozrejme, vysloviť hypotéza, že slovenské smrečiny majú „boreálny charakter“ a teda sa svojim vývojovým cyklom líšia od „bežných“ lesov mierneho pásma. Aké sú teda argumenty pre alebo proti tejto teórii?
V boreálnych lesoch je jedným z dôkazov veľkého cyklu hojný veľkoplošný výskyt pionierskych drevín. U nás sú, naproti tomu, rozsiahlejšie porasty pionierskych drevín vzácne, a to aj na neprístupných lokalitách, kde tento stavu pravdepodobne nie je výsledkom hospodárenia. V prípade, že sa takéto porasty miestami vo väčšom počte vyskytujú, je zvyčajne známa antropogénna príčina ich vzniku (napr. požiar). Najbližšie k drevinovému zloženiu boreálnych lesov majú porasty juhovýchodných svahov Vysokých Tatier (v súčasnosti z veľkej časti zničené kalamitou z r. 2004), v ktorých zvýšené zastúpenie smrekovca a borovice svedčí o častých disturbanciách. Nedá sa však povedať, či šlo o disturbancie prirodzené, alebo len o dôsledok zmeny štruktúry týchto lesov človekom.

Prírodná rezervácia Fabova hoľa (stav v septembri 2008)
Ponechanie menších vetrových kalamít spôsobilo masívny rozpad smrečín, ktorých prirodzenosť je navyše v spodnej časti rezervácie (hranicou je lesná cesta viditeľná na fotografii) sporná. Rezervácia je obklopená prevažne smrekovými hospodárskymi lesmi, ktoré už sú v súčasnosti napádané podkôrnym hmyzom.
Foto: Ing. Matej Schwarz
Navyše treba zdôrazniť, že veľký cyklus vo svojej typickej podobe nie je, a nikdy nebol, spôsobený aktivitou podkôrneho hmyzu (pozri vyššie). Pokiaľ by sa totiž podkôrny hmyz stal hnacou silou tohto cyklu, nevyhnutne by sa to prejavilo znížením dosiahnuteľného veku smrečín. Napr. lykožrút smrekový napáda porasty nad cca 60 rokov, takže postupne by sa staršie porasty prestali v jeho areáli vyskytovať. Keďže takto mladé porasty nebývajú plne plodivé, pravdepodobne by časom nastali problémy s obnovou smreka. Okrem toho sú známe prípady, keď lykožrút smrekový po „spotrebovaní“ dospelých smrekov začal napádať aj smrekové mladiny. Mladé porasty sú napádané aj niektorými ďalšími (hoci menej nebezpečnými) druhmi podkôrneho hmyzu. Významná je aj skutočnosť, že podkôrny hmyz by sa stal takmer výlučným faktorom prirodzeného výberu, takže by stromy prestali byť selektované na odolnosť voči vetru, snehu a p., čím by sa u nich pravdepodobnosť dožívania sa vysokého veku znížila aj v prípade, že by tlak podkôrneho hmyzu v budúcich desaťročiach ustal. Všetky tieto okolnosti by postupne viedli k premene smrečín na iný typ biotopu.
Okrem toho jednoduché simulácie dokazujú, že masové odumieranie lesa, ktoré je typické pre súčasné pôsobenie podkôrneho hmyzu, nevyhnutne vedie k vzniku rozsiahlych rovnovekých porastov. Tento hmyz je totiž pre premnožení ničiť stromy od spomenutých 60 rokov až po najstaršie. Takto je v krátkom čase 8 až 12 vekových tried nahradených jedinou.
Celkovo teda možno konštatovať, že stav našich smrečín nenasvedčuje tomu, že by v nich v minulosti masovo prebiehal veľký vývojový cyklus a vôbec už nie tomu, že by tento cyklus býval riadený podkôrnym hmyzom. Aj keď sa nedá vylúčiť, že tento hmyz hral aj v pôvodných lesoch určitú úlohu, jeho pôsobenie sa nemohlo ani len blížiť jeho dnešnému vplyvu. Existencia starých smrekových porastov (bežne až 300 ročných), alebo rôznovekých smrečín s dostatočným počtom starých stromov dokazujú, že po dobu takmer sa rovnajúcu ich veku sa takto závažné premnoženia podkôrneho hmyzu v ich okolí nevyskytli.
Bez ohľadu na možnú prirodzenosť veľkého cyklu v jednotlivých porastoch je zo stavu našich smrečín zrejmé, že k nemu prinajmenšom nedochádzalo naraz na veľkých plochách. Je teda možné povedať, že samovoľná premena jednotlivého smrekového porastu na porast pionierskych drevín je prirodzená, z toho však nijako nevyplýva, že to isté by malo byť prirodzené v rozsahu tisícov hektárov. Navyše, aj v prípade, že takýto vývoj by v minulosti býval prirodzený, v súčasnosti nie je príliš akceptovateľný. V dnešnej krajine s prerušenými migračnými koridormi a nedostatkom pôvodného genofondu, by masové odumieranie porastov viedlo k rýchlej degradácii chránených území resp. lesov všeobecne.
Pokiaľ majú byť chránené územia trvalo udržateľné, výmera nižších vývojových štádií lesa nesmie presiahnuť podiel primeraný dĺžke celého vývojového cyklu. Toto platí rovnako pre dynamiku hospodárskeho lesa, ako pre dynamiku prirodzenú. Ak sa napr. najstaršie porasty majú dožiť 200 rokov, ročne je možné vyrúbať len 0,5 % celkovej výmery územia. Ak by sme ročne rúbali napr. 1%, les by bol rúbaný rýchlejšie ako stačí dorastať. V prípade, že namiesto rúbania dochádza k rozpadu porastov pôsobením podkôrneho hmyzu, je situácia zložitejšia, pretože pre návrat k pôvodnému stavu môže byť potrebných viacero sukcesných štádií. Ak napr. vývoj vedie cez štádium „mŕtveho lesa“ trvajúceho 10 rokov, jarabinového lesa trvajúceho 50 rokov a až potom sa začne vývoj smrečiny, celý cyklus sa predĺži na 260 rokov a ročne teda smie odumrieť len 0,38%. Pokiaľ by to bolo viac, dosiahnuteľný vek a výmera klimaxového sukcesného štádia, budú klesať. Výmera nižších vývojových štádií je teda významným indikátorom trvalej udržateľnosti chráneného biotopu.
[5] Napríklad Míchal, I. a kol. 1994: Ekologická stabilita, Veronica.